CogSci 30PY ProjectThe IT Cortex |
"The brain is not an organ of thinking but an organ of survival, like claws and fangs. It is made in such a way as to make us accept as truth that which is only advantage. It is an exceptional, almost pathological constitution one has, if one follows thoughts logically through, regardless of consequences. Such people make martyrs, apostles, or scientists, and mostly end on the stake, or in a chair, electric or academic." -Albert Szent-Gyorgi |
![]()
http://www.comp.leeds.ac.uk/ugadmit/cogsci/tech/hopn.htm:
Hopfield nets can remeber 0.14N states stably, where N is the number
of units in the net.
http://www.ida.his.se/ida/kurser/ai_ann/kursmaterial/archive/lectureA.pdf:
Synchronous updates may lead to oscillation and the network may not always
converge.
Is this what we need for binocular rivalry?
http://www.nada.kth.se/~asa/hopfield.ps:
for synchronous updating the network may end up flipping between two (but
not more) states.
--------------------------------------------------------------------------------------
David J.C. MacKay
Information Theory, Inference, and Learning Algorithms
Ch 32/33 Exact Monte Carlo Sampling / Variational Methods Ch 42 references 33.2, but it is all maths!
Ch 39 The Single Neuron as classifier
Ch 40 Capacity of a Single Neuron
Ch 41 Learning as inference
Ch 42 Hopfield Networks - Explanation starts with Hebbian learning -> Hopfield NNs!
Ch 43 Boltzmann Machines
Hopfield Networks / Boltzmann machines
This book is also available from Amazon
-------------------------------------------------------------------------
C
Files for the book "An Introduction to Neural Networks"
by
James A. Anderson,
Department of Cognitive and Linguistic Sciences,
Brown University, Providence, RI 02912., on MIT Press.
-------------------------------------------------------------------------
Hopfield
example code from "Neural Networks at Your Fingertips"
--------------------------------------------------------------------------
Hopfield Networks at Wolfram Research:
Hopfield networks, or associative networks, are typically used for classification. Given a distorted input vector, the Hopfield network associates it with an undistorted pattern stored in the network.
--------------------------------------------------------
NASATech says: The second architecture is that of autoassociative neural networks, which, in nature, enable organisms to recall old memories from partial or noisy stimuli. In an autoassociative network, each neuron is connected to every other neuron in the network. The excitability of any such neuron is determined by the state of all the other neurons and the strengths (weights) of the interconnections between neurons. For any state, the pattern of activation at the next instant in time is completely determined by the preset weights. The number of different firing patterns is 2^n, where n is the number of neurons in the net; because this number is finite, as the sequence of firing patterns proceeds, cycles are inevitable. Cycles that consist of repeated instances of the same pattern are called fixed points, and these fixed points can be chosen by setting the weights to make the points attract nearby patterns. The fixed points represent memories. An autoassociative sensor web could be used to search for known gaseous, biological, or geological signatures, for example.
A chapter on Attractor Networks. This is a good explanation of some of the terms (like "attractor basin").
Oxford Lecture notes on Hopfield Networks and Alzheimer's. Here's a cut from it:

Fusi's PhD
thesis, which I've linked to before, seems relevant:
It is about the IT
cortex;
context-dependent
memory;
recognition but also a no-recognition state (i.e.,
a spontaneous activity state)
-- p. 6, bottom.
Following are some extracts from his thesis.
From the thesis. Page numbers are from the pdf, not the printed numbers on the page. (My italics):
Another key property of slow learning is that memory of the class prototypes is always stronger than memory of any other members of the same class which have been shown. This is essential for the classification capability and for reproducing the behavior of the paradigmatic attractor cell. (p. 11)
...the process of learning can be fully described
in terms of a few parameters which are the
transition probabilities for LTP and LTD. (p. 11)
...learning requires many presentations of the same stimulus... (p. 11) (over 500)
Stochastic learning provides a good tool for studying
quantitatively the process of attractor
formation without specifying the details of the synaptic dynamics. However,
to proceed
further, it is important to have a framework that provides a link between
the transition
probabilities and the neural activity driven synaptic dynamics. This implies
the identification of the source of noise which drives the stochastic
mechanism. The solution we propose is suggested by the analysis of cortical
recordings: the spike trains recorded in vivo are similar to poissonian
processes [Softky Koch 1993] and the variability in the intervals between
the spikes of pre- and post-synaptic neurons is a quantity that can be
\read" by the synapses. What we show in chapter 7.1 is that it is
possible to use this source of
stochasticity to obtain transitions with small probabilities. (p. 13)
Persistent enhanced spike rates have been observed
throughout the delay interval between
two successive visual stimulations in several cortical areas: e.g. they
are encountered in
infero-temporal (IT) cortex [Fuster, 1990, Fuster & Jervey, 1981,
Miyashita & Chang, 1988,
Miyashita, 1988, Sakai & Miyashita, 1991, Miller et al, 1993, Nakamura
& Kubota, 1995],
in pre-frontal (PF) cortex [Funahashi et al, 1989, Wilson et al. 1993,
Miller et al, 1996,
Scalaidhe et al, 1997, Rao et al. 1997] and in posterior parietal cortex
[Koch &Fuster, 1989]
(for a review see [Fuster, 1995]). The phenomenon is detected in single
unit extra-cellular
recordings of spikes following, rather than during, the presentation of
a sensory stimulus. It
has been found in primates trained to perform a delayed response task,
in which the behav-
ing monkey must remember the identity or location of an initial eliciting
stimulus in order
to decide upon its behavioral response. The pattern of delay activity
is considered a neural
correlate of working memory, which is related to the ability of the
animal to actively hold an
item in memory for a short time. In fact, lesions in IT and PF are known
to produce impair-
ments in spatial and associative memory tasks [Murrayet al. 1993, Gutnikov
et al. 1997],
and a ect recognition of an object after a delay in both humans and monkeys.
(p. 15)
The collective aspect is expressed in the mechanism
by which a stimulus that had
been learned previously, has left a synaptic engram of potentiated
excitatory synapses
connecting the cells driven by the stimulus. When this subset of cells
is re-activated by
a sensory event, they cooperate to maintain elevated ring rates in the
selective set of
neurons, via the same set of potentiated synapses, after the stimulus
is removed. In this
way, these cells can provide each other, i.e. each one of the group, with
an a afferent signal
that clearly differentiates the members of the group from other cells
in the same cortical
module. (p. 18)
Attractor dynamics (associative memory) implies that each pattern of
delay activity is
excited by a whole class of stimuli. Each of these stimuli raises the
rates of a sufficient
number of cells belonging to the pattern of activity, so that via the
learned collateral
synaptic matrix they can excite the entire subset of neurons characterizing
the attractor
and maintain them with elevated rates when the stimulus is removed. (p.
25)
The class of stimuli leading to the same persistent distribution is the
basin of attraction of that particular attractor. (pp. 25-26) As one moves
in the space of stimuli, at some point the boundary of the basin
of attraction of the attractor is reached. Moving even slightly beyond
this boundary,
the network will relax either into another learned attractor, or to the
grand attractor of
spontaneous activity. See e.g. [Amit & Brunel, 1997a, Amit, 1995].
(p. 26)
The dynamics of the simplest integrate-and-fire (IF) neuron can
be fully described in terms
of a single internal variable: the depolarization V (t) of the neural
membrane. The neuron
integrates the afferent current, and when the depolarization crosses a
threshold, the neuron
emits a spike. The depolarization is then reset to the hyperpolarization
value H, and the
integration process starts again after an absolute refractory period Tarp.
Formally, the differential equation which governs the dynamics below threshold,
can be
written as follows:
dV (t) / dt = -L(t1V) + I(t),
where I(t) is the net charging current, expressed in units of potential
per unit time, produced by afferent spikes, and L(t) is the leakage term.
Usually it is proportional to the depolarization: L(t) = -V(t)/r, where
r is the integration time constant of the membrane depolarization. A linear
constant decay, not dependent
on V, L(t) = B simplifies the analysis of the collective behavior of the
network without sacrificing those features that allow to reproduce the
phenomenology of cortical recordings [Fusi & Mattia 1999].
(p. 31. has properly formatted equations)
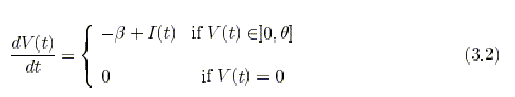 (p. 32)
(p. 32)
This neuron he calls the LIF neuron (Linear Integrate-and-Fire neuron).3.1.1 The anatomy of the network module
We consider a local module that is a large network containing a number
of neurons of
the order of 10^4 - 10^5 . The number of connections between neurons of
the same module is
higher than the number of connections between neurons belonging to different
modules.
Each neuron is randomly connected to a subset of neurons of the same module
through the
recurrent collaterals, and to neurons of other modules through longer
range connections
(see Fig.3.1).
In particular, the network module is composed of excitatory (pyramidal)
and inhibitory
(inter-neurons) neurons. Each neuron has some probability (connectivity
level) of a
synaptic contact with any other neuron (excitatory or inhibitory) of the
same module, and
receives external excitatory afferents from pyramidal neurons of other
modules.
The connectivity structure can be fully described in terms of two sets
of variables:
the binary variables cij of having a connection between neuron j (pre-synaptic
neurons)
and neuron j (post-synaptic neuron) and the corresponding synaptic weight
Jij . In what
follows the cij are generated at random and never changed, while the synaptic
couplings
Jij can vary to acquire information about the stimuli that excite part
of the neurons of the
network module. (p. 32)
SD = signal dominated (p. 33)
ND = noise dominated (p. 34)
Each neuron will receive a current Ii(t) (i denotes the index of the neuron) given by:
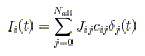 (p. 32)
(p. 32)
where cij is 1 if there exists a synaptic contact between neuron j and
neuron i, Jij is
the corresponding synaptic strength, and the sum extends over all the
Nall neurons of the
populations (recurrent or external) that are interacting.
"Such being the nature of our subject and such our way of arguing in our discussion of it, we must be satisfied with a rough outline of the truth."
-Aristotle
![]()